
COURTSHIP BEHAVIOR
| main |
| behavior |
| pheromone |
| receptors |
| neural response |
| publications |
| people |
|
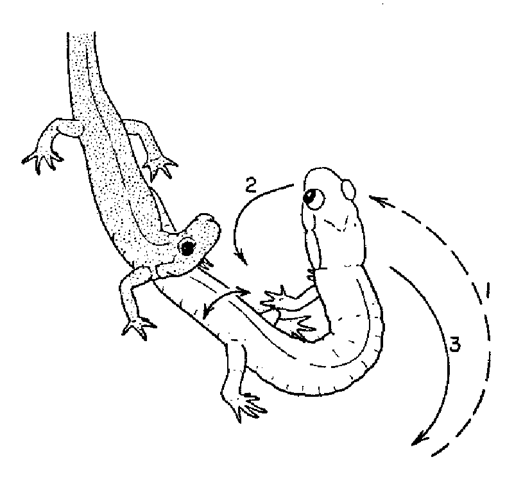
During courtship, a male P. shermani delivers pheromones to the female's VNO by slapping (2) his mental gland across her nares. (Arnold 1976).
|





| Courtship Behavior & Female Sexual Receptivity |
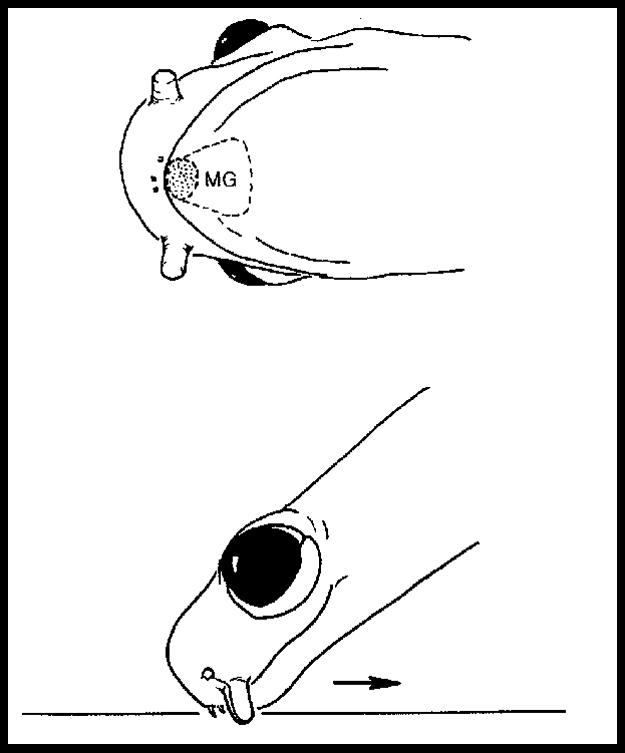
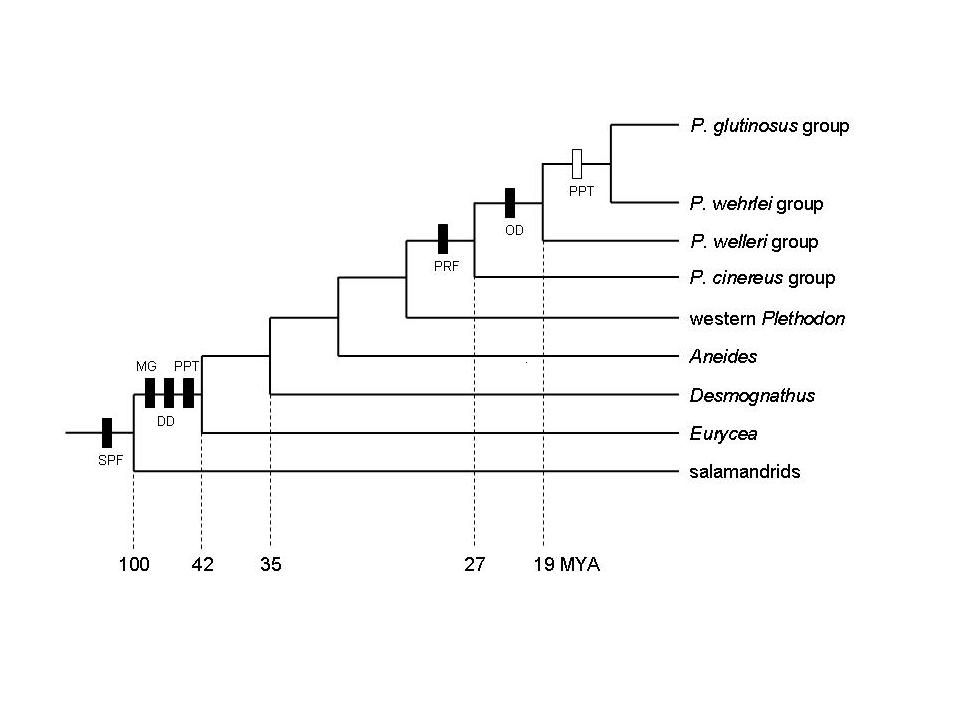
Much of our behavioral work has focused on Plethodon shermani (formerly P. jordani), a plethodontid salamander with complex, stereotyped courtship behavior (Arnold 1976, Houck 1986, Marvin & Hutchinson 1996, Picard 2005, Dyal 2006). Stage 1. Orientation. The male orients to the female and moves towards her. PHEROMONE EFFECTS ON FEMALE BEHAVIOR During courtship, a male P. shermani salamander delivers pheromones from a submandibular mental gland to the female’s nares. The pheromones are only delivered after courtship has commenced and their purpose is to increase female receptivity and mating success. These pheromones do not serve as a sexual attractant. We conducted behavioral tests in which the natural delivery of male courtship pheromones was prevented by ablating the courtship (mental) glands of all experimental males in order to control experimentally the amount and type of pheromone stimuli received by each female. We conducted three behavioral tests, each using our standard protocol. In our first test, we used a pheromone solution that was enriched (relative to untreated gland extracts) with the protein component PRF (i.e., PRF accounted for approx. 90% of proteins in this solution). For courting pairs in which the female received this enriched solution, female receptivity was increased (i.e., courtships were completed more quickly) (Houck et al. 1998). In our second behavioral experiment, we similarly staged courtship encounters which demonstrated that a PRF solution that was purified (i.e., no other detectable proteins were present) was effective in increasing female receptivity (Rollmann, Houck & Feldhoff, 1999). In our third experiment, we demonstrated that female response to courtship pheromones from conspecific sympatric males and from heterospecific allopatric males was statistically indistinguishable (Rollmann, Houck & Feldhoff 2003). EVOLUTIONARY STASIS IN MALE COURTSHIP BEHAVIOR AND MORPHOLOGY The pheromone delivery system of plethodontid salamanders presents a remarkable picture of morphological and behavioral stasis (Houck & Sever 1994, Houck & Arnold 2003). Species in each of the four major lineages share the same, complex system of pheromone delivery by vaccination. During the courtship season, males grow hypertrophied premaxillary teeth and develop a small pad of glandular tissue on their chins (the mental gland). During courtship, the male abrades the female's skin with his premaxillary teeth and rubs secretions from his mental gland into the abraded site. These secretions increase female receptivity and shorten the time to sperm transfer (Houck & Reagan 1990). Because this complex system is found in all four major lineages of plethodontids and in no other salamander family, it was undoubtedly present in the ancestral plethodontid. Thus, because the family itself is approximately 100 million years old (Ruben et al. 1993), the morphological and behavioral elements in the delivery system have been conserved over that entire period. Charlesworth et al. (1982) have persuasively argued that such long term stasis must be a consequence of stabilizing selection. Other mechanisms – such as evolutionary inertia or developmental constraint – may produce short term stasis, but they cannot account for long term stasis. An equally puzzling example of stasis is found within the genus Plethodon. The forty-one species of this genus found in eastern North America form a monophyletic group with two subdivisions that differ greatly in body size: the small eastern Plethodon and the large eastern Plethodon (Highton & Larson 1979, Larson et al. 1981, Highton 1993, Highton & Peabody 2000). The small eastern Plethodon (12 species) retain the ancestral mode of vaccination delivery of pheromones. The large eastern species (29 species) are a monophyletic group that share an olfactory mode of pheromone delivery. In these species, the males lack protruding premaxillary teeth and possess a large circular mental gland. These species deliver pheromones by slapping their large mental glands over the female's snout during courtship. Immunological and allozyme data indicate that the radiation of large eastern Plethodon – and hence the olfactory mode of delivery – is about 19 million years old (Highton & Larson 1979). Although we do not understand the details of the process that is responsible for this second example of stasis, under the same argument as before, it is probably a consequence of stabilizing selection maintained over a 19 million year period. |